ARG56158
anti-PLK1 antibody
anti-PLK1 antibody for ICC/IF,IHC-Formalin-fixed paraffin-embedded sections,Western blot and Human,Mouse,Rat

1
Overview
| Product Description | Rabbit Polyclonal antibody recognizes PLK1 |
|---|---|
| Tested Reactivity | Hu, Ms, Rat |
| Tested Application | ICC/IF, IHC-P, WB |
| Host | Rabbit |
| Clonality | Polyclonal |
| Isotype | IgG |
| Target Name | PLK1 |
| Antigen Species | Human |
| Immunogen | Recombinant protein of Human PLK1. |
| Conjugation | Un-conjugated |
| Alternate Names | STPK13; Serine/threonine-protein kinase PLK1; EC 2.7.11.21; Serine/threonine-protein kinase 13; Polo-like kinase 1; PLK-1; PLK |
Application Instructions
| Application Suggestion |
|
||||||||
|---|---|---|---|---|---|---|---|---|---|
| Application Note | * The dilutions indicate recommended starting dilutions and the optimal dilutions or concentrations should be determined by the scientist. |
Properties
| Form | Liquid |
|---|---|
| Purification | Affinity purification with immunogen. |
| Buffer | PBS (pH 7.3), 0.02% Sodium azide and 50% Glycerol. |
| Preservative | 0.02% Sodium azide |
| Stabilizer | 50% Glycerol |
| Storage Instruction | For continuous use, store undiluted antibody at 2-8°C for up to a week. For long-term storage, aliquot and store at -20°C. Storage in frost free freezers is not recommended. Avoid repeated freeze/thaw cycles. Suggest spin the vial prior to opening. The antibody solution should be gently mixed before use. |
| Note | For laboratory research only, not for drug, diagnostic or other use. |
Bioinformation
| Database Links | |
|---|---|
| Gene Symbol | PLK1 |
| Gene Full Name | polo-like kinase 1 |
| Background | The Ser/Thr protein kinase encoded by this gene belongs to the CDC5/Polo subfamily. It is highly expressed during mitosis and elevated levels are found in many different types of cancer. Depletion of this protein in cancer cells dramatically inhibited cell proliferation and induced apoptosis; hence, it is a target for cancer therapy. [provided by RefSeq, Sep 2015] |
| Function | Serine/threonine-protein kinase that performs several important functions throughout M phase of the cell cycle, including the regulation of centrosome maturation and spindle assembly, the removal of cohesins from chromosome arms, the inactivation of anaphase-promoting complex/cyclosome (APC/C) inhibitors, and the regulation of mitotic exit and cytokinesis. Polo-like kinase proteins acts by binding and phosphorylating proteins are that already phosphorylated on a specific motif recognized by the POLO box domains. Phosphorylates BORA, BUB1B/BUBR1, CCNB1, CDC25C, CEP55, ECT2, ERCC6L, FBXO5/EMI1, FOXM1, KIF20A/MKLP2, CENPU, NEDD1, NINL, NPM1, NUDC, PKMYT1/MYT1, KIZ, PPP1R12A/MYPT1, PRC1, RACGAP1/CYK4, SGOL1, STAG2/SA2, TEX14, TOPORS, p73/TP73, TPT1 and WEE1. Plays a key role in centrosome functions and the assembly of bipolar spindles by phosphorylating KIZ, NEDD1 and NINL. NEDD1 phosphorylation promotes subsequent targeting of the gamma-tubulin ring complex (gTuRC) to the centrosome, an important step for spindle formation. Phosphorylation of NINL component of the centrosome leads to NINL dissociation from other centrosomal proteins. Involved in mitosis exit and cytokinesis by phosphorylating CEP55, ECT2, KIF20A/MKLP2, CENPU, PRC1 and RACGAP1. Recruited at the central spindle by phosphorylating and docking PRC1 and KIF20A/MKLP2; creates its own docking sites on PRC1 and KIF20A/MKLP2 by mediating phosphorylation of sites subsequently recognized by the POLO box domains. Phosphorylates RACGAP1, thereby creating a docking site for the Rho GTP exchange factor ECT2 that is essential for the cleavage furrow formation. Promotes the central spindle recruitment of ECT2. Plays a central role in G2/M transition of mitotic cell cycle by phosphorylating CCNB1, CDC25C, FOXM1, CENPU, PKMYT1/MYT1, PPP1R12A/MYPT1 and WEE1. Part of a regulatory circuit that promotes the activation of CDK1 by phosphorylating the positive regulator CDC25C and inhibiting the negative regulators WEE1 and PKMYT1/MYT1. Also acts by mediating phosphorylation of cyclin-B1 (CCNB1) on centrosomes in prophase. Phosphorylates FOXM1, a key mitotic transcription regulator, leading to enhance FOXM1 transcriptional activity. Involved in kinetochore functions and sister chromatid cohesion by phosphorylating BUB1B/BUBR1, FBXO5/EMI1 and STAG2/SA2. PLK1 is high on non-attached kinetochores suggesting a role of PLK1 in kinetochore attachment or in spindle assembly checkpoint (SAC) regulation. Required for kinetochore localization of BUB1B. Regulates the dissociation of cohesin from chromosomes by phosphorylating cohesin subunits such as STAG2/SA2. Phosphorylates SGOL1: required for spindle pole localization of isoform 3 of SGOL1 and plays a role in regulating its centriole cohesion function. Mediates phosphorylation of FBXO5/EMI1, a negative regulator of the APC/C complex during prophase, leading to FBXO5/EMI1 ubiquitination and degradation by the proteasome. Acts as a negative regulator of p53 family members: phosphorylates TOPORS, leading to inhibit the sumoylation of p53/TP53 and simultaneously enhance the ubiquitination and subsequent degradation of p53/TP53. Phosphorylates the transactivation domain of the transcription factor p73/TP73, leading to inhibit p73/TP73-mediated transcriptional activation and pro-apoptotic functions. Phosphorylates BORA, and thereby promotes the degradation of BORA. Contributes to the regulation of AURKA function. Also required for recovery after DNA damage checkpoint and entry into mitosis. Phosphorylates MISP, leading to stabilization of cortical and astral microtubule attachments required for proper spindle positioning. Together with MEIKIN, acts as a regulator of kinetochore function during meiosis I: required both for mono-orientation of kinetochores on sister chromosomes and protection of centromeric cohesin from separase-mediated cleavage (By similarity). [UniProt] |
| Calculated MW | 68 kDa |
| PTM | Catalytic activity is enhanced by phosphorylation of Thr-210. Phosphorylation at Thr-210 is first detected on centrosomes in the G2 phase of the cell cycle, peaks in prometaphase and gradually disappears from centrosomes during anaphase. Dephosphorylation at Thr-210 at centrosomes is probably mediated by protein phosphatase 1C (PP1C), via interaction with PPP1R12A/MYPT1. Autophosphorylation and phosphorylation of Ser-137 may not be significant for the activation of PLK1 during mitosis, but may enhance catalytic activity during recovery after DNA damage checkpoint. Phosphorylated in vitro by STK10. Ubiquitinated by the anaphase promoting complex/cyclosome (APC/C) in anaphase and following DNA damage, leading to its degradation by the proteasome. Ubiquitination is mediated via its interaction with FZR1/CDH1. Ubiquitination and subsequent degradation prevents entry into mitosis and is essential to maintain an efficient G2 DNA damage checkpoint. Monoubiquitination at Lys-492 by the BCR(KLHL22) ubiquitin ligase complex does not lead to degradation: it promotes PLK1 dissociation from phosphoreceptor proteins and subsequent removal from kinetochores, allowing silencing of the spindle assembly checkpoint (SAC) and chromosome segregation. |
Images (1) Click the Picture to Zoom In
-
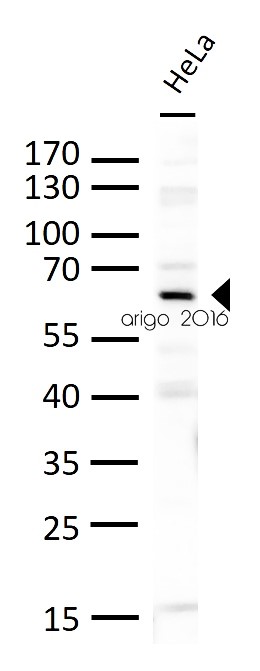
ARG56158 anti-PLK1 antibody WB image
Western blot: 30 µg of HeLa cell lysates stained with ARG56158 anti-PLK1 antibody at 1:500 dilution.
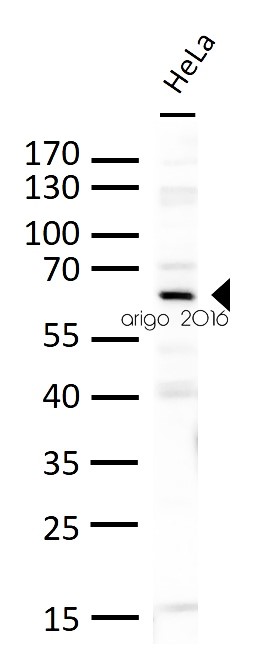
Customer's Feedback
Good
Review for anti-PLK1 antibody
Application:WB
Sample:HeLa
Sample Loading Amount:30 µg
Primary Antibody Dilution Factor:1:500
Primary Antibody Incubation Time:overnight
Primary Antibody Incubation Temperature:4 ºC